
Evidence suggests that SARS-CoV-2, as well as other coronaviruses, can be dispersed and potentially transmitted by aerosols directly or via ventilation systems. We therefore investigated ventilation openings in one COVID-19 ward and central ducts that expel indoor air from three COVID-19 wards at Uppsala University Hospital, Sweden, during April and May 2020. Swab samples were taken from individual ceiling ventilation openings and surfaces in central ducts. Samples were subsequently subjected to rRT-PCR targeting the N and E genes of SARS-CoV-2. Central ventilation HEPA filters, located several stories above the wards, were removed and portions analyzed in the same manner. In two subsequent samplings, SARS-CoV-2 N and E genes were detected in seven and four out of 19 room vents, respectively. Central ventilation HEPA exhaust filters from the ward were found positive for both genes in three samples. Corresponding filters from two other, adjacent COVID-19 wards were also found positive. Infective ability of the samples was assessed by inoculation of susceptible cell cultures but could not be determined in these experiments. Detection of SARS-CoV-2 in central ventilation systems, distant from patient areas, indicate that virus can be transported long distances and that droplet transmission alone cannot reasonably explain this, especially considering the relatively low air change rates in these wards. Airborne transmission of SARS-CoV-2 must be taken into consideration for preventive measures.
Introduction
During the coronavirus infectious disease 19 (COVID-19) pandemic, droplet transmission has been considered the most significant transmission route for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), although other routes such as aerosol, fecal–oral, and indirect transmission via fomites may contribute to the rapid global dissemination of the virus1,2. The relative importance of aerosols versus droplets in the transmission of respiratory infections is difficult to distinguish, since particles of both aerosol and droplet size are generated for example when talking3,4. Aerosols are smaller than droplets, traditionally defined as smaller than 5 µm in diameter, and are thought to remain airborne longer, enabling transmission at greater distances and over longer periods of time5. This definition has been challenged and may very well be an over-simplification and it may be precarious to rigidly differentiate the two categories3,6,7.
Previously, other coronaviruses have been shown to disperse via aerosols and ventilation, and have been determined to cause HVAC (heating, ventilation, air conditioning) associated and nosocomial infections as well as extensive hospital outbreaks8,9,10,11,12,13. In recent studies, extensive environmental contamination of SARS-CoV-2 in hospital settings has been demonstrated, and viral RNA has been found both in air samples and in samples from air vent openings in isolation rooms14,15,16,17,18. Also, the potential for the aerosol transmission route of SARS-CoV-2 is supported by other recent studies17,19,20,21. The increased risk for infection in indoor environments, as well as superspreading events, could be explained by airborne transmission22,23,24,25,26. In this context it is therefore vital to understand the amount of SARS-CoV-2 in confined spaces and the distances at which virus can be passively dispersed. Hospital rooms where COVID-19 patients are treated are obviously venues in which airborne transmission is both of great importance to understand, as well as a suitable environment to study this phenomenon. In this study from a COVID-19 infectious disease ward at Uppsala University Hospital, Sweden, we investigated if SARS-CoV-2 RNA could be detected in and near air vent openings in isolation rooms and in filters in the central ventilation system situated on the eighth (top) floor of the hospital building. As RNA was detected at substantial distances from patient areas, fluid sample collections were performed in an attempt to determine the potential infective ability of SARS-CoV-2 detected in the systems. Our findings may suggest both airborne dispersal of SARS-CoV-2 and possible long-distance dissemination of SARS-CoV-2 via ventilation air flow.
Materials and methods
Sampling strategy
Sampling was performed on separate occasions during April and May 2020. In the first two occasions, 17 and 28 April, surfaces of exit vent openings in all 19 patient rooms in ward 1 (Fig. 1a) were swabbed as described below. When repeated on April 28, the internal surfaces of the central ventilation ducts, on the top floor were also swabbed and filter sections removed, as described further below. Due to the detection of SARS-CoV-2 RNA in the ventilation system (see “Results”), a further sample collection was performed using fluid traps, both at the terminal end of the ducts prior to the exhaust filters (at the same area where swabs were taken on April 28) as well as under the ceiling vent openings in the ward rooms (ward 1, see Fig. 1b), in an attempt to determine the infective ability of any collected virus.
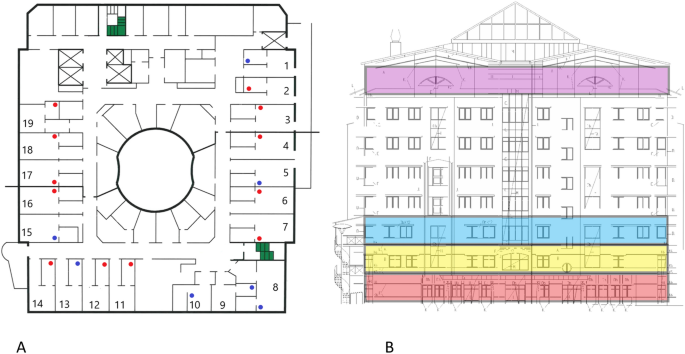
(A) Overview of the 19 investigated COVID-19 ward rooms (ward 1). Dots indicate approximate placing of ceiling vent openings. Red dots indicate openings that where SARS-CoV-2 RNA was detected in at least one of two samplings, blue dots openings negative in both samplings. (B) Lateral view of the hospital building. Ward levels: red; COVID-19 outpatient clinic, yellow and blue; COVID-19 wards 1 and 2, with 19 rooms each, purple; eighth floor with central ventilation fans and HEPA filters. Individual ceiling vent openings were investigated on the second-floor ward (yellow) seen in (A).
Swab samples
Surfaces were swabbed using sterile nylon flocked swabs (Copan eSwab, Copan Italia SpA, Italy) moistened in sterile viral transport medium (VTM), containing Hank’s balanced salt solution (Gibco, UK) supplemented with 2% fetal bovine serum (Gibco, USA), 100 µg/ml Gentamicin, and 0.5 µg/ml Amphotericin B27. Round ceiling vent openings were swabbed around the inside of the entire opening (circumference ca 25 cm). Swabs were placed in tubes containing 750 µl viral transport medium and stored at 4 °C until analysis within 24–72 h. Sampling was performed on April 17 and 28, 2020. Indoor relative air humidity and temperature were 30–31% and 20–21 °C, respectively.
Filter samples
Exit ventilations from each of the eight stories in the investigated hospital building, (Fig. 1b), lead to separate HEPA filter systems, located on the eighth (top) floor. Consequently, we could identify ducts and exhaust filters collecting air from individual floors not merging airflows. We chose to examine exhaust filters from three floors in the building that had been specifically designated for COVID-19 patients; two COVID-19 wards and a COVID-19 out-patient clinic. In addition, we examined exhaust filters from one story with personnel areas and a cafeteria, as a negative control. The distance between the COVID-19 wards and the exhaust filters and inspection hatches was 49, 53 and 56 m respectively for each COVID-19 ward (Table 1). The four stories located between the COVID-19 wards and the central ventilation in the top of the building (Fig. 1b) only sporadically harbored COVID-19 patients and were therefore not investigated. Adjacent inspection hatches upstream from (prior to) the HEPA filters were opened, and internal 30 × 30 cm surfaces swabbed as described above. Furthermore, one (out of six) 60 × 60 cm laminate F7 HEPA filter sections was removed from each system (filtering air from one ward or floor) and three filter samples (3 × 3 cm) were randomly cut out of the filters using sterilized scissors, placed in vials containing 2.5 ml of viral transport medium (described above), and stored at 4 °C until analysis within 72 h. The removed filters had been routinely replaced one month prior to collection.
Fluid samples
Fluid sample collection was performed near air entrances (ward rooms) and exits (directly prior to exhaust filters) in the ventilation system by placing open, 10 cm diameter petri dishes with 10 ml of DMEM (Dulbecco’s Modified Eagle’s medium (cell medium); Gibco) diluted 1 to 5 with autoclaved water, suspended 15 cm below ceiling vent openings (in ward rooms) for 24 h, or placed within central vent ducts via inspection hatches for 3 h. DMEM was diluted to ensure appropriate salt balance for the cells and no osmotic effect on the virus after evaporation of water during the collection process. We used DMEM instead of water only to be able to add the whole volume of sample onto cells without a dilution effect of the cell medium. These points were chosen in an attempt to determine if virus found entering and/or exiting the ventilation ducts retained infective ability, in response to the PCR results from vent opening and exhaust filters. The suspended petri dishes in the ward rooms were placed within what we expected to be the normal air flow to ventilation ducts, as well as placing the dishes as far from the patients as possible, in order to avoid contamination by coughing induced droplets or other patient or personnel activity in the rooms. An open petri dish containing cell medium was exposed to air in the biosafety level (BSL)-2 area of the laboratory for 24 h and used as a negative control, along with non-exposed DMEM and viral transport medium. DMEM and VTM spiked with synthetic oligonucleotides (gBlocks, IDT, Belgium) based on N and E gene sequences with introduced 5 base pair deletions were used as positive controls (Suppl. Table 1). DMEM exposed to air in 19 ward rooms were combined to three pools. Pooling was performed when we could establish that a large number of rooms were occupied by non-contagious patients (seven rooms), patients with suspected COVID-19 but not confirmed (five rooms) and only six rooms were occupied by contagious COVID-19 patients (May 13, 2020) (Suppl. Table 1). Due to evaporation during collection, the final concentration of DMEM in the petri dishes after collection was equivalent to undiluted cell medium. The entire pooled volume ranging from 5 to 10 ml was subsequently applied to Vero E6 cells in T25 flasks and incubated up to 13 days. Samples were subsequently collected and subject to rRT-PCR. Petri dishes with 10 ml DMEM exposed to air outside of patient areas for 24 h were used as negative control.
Ward conditions
All exit vent openings in the ward rooms are situated in the ceiling and are approximately 3 to 5 m from the head end of the beds (fresh air input openings are at 0 to 50 cm above floor level). Seven of the 19 openings are situated in adjacent washing rooms (see Fig. 1a) and are up to approximately 5 to 6 m from beds. Total air changes per hour (ACH) for each patient room varied between 1.5 and 2.6 in ward 1, and 2.1 to 2.7 in ward 2, between 2.8 and 3.2 in the outpatient clinic, (measured December 2017). Air flow in the central ventilation shafts, from each story, ranged between 2.27 and 3.48 m3/s (Table 1). Pressure differences in rooms in ward 1 varied, − 6 to − 8.1 Pa between corridor and anterooms and + 5.5 to + 18 Pa between anterooms and patient rooms (measured March 2020). Hence, the anterooms were under negative pressure compared to the adjacent ward corridor as well as patient rooms.
RNA extraction and rRT-PCR
RNA was extracted using 280 μl of samples and QIAamp viral RNA kit (Qiagen, Hilden, Germany), according to manufacturer’s protocol. Portions of the SARS-CoV-2 nucleocapsid (N) and envelope small membrane protein (E) genes were amplified by rRT-PCR, using primers (Thermo Fisher Scientific, Waltham, MA, USA) previously described28,29,30 and the SuperScript III OneStep RT-PCR System with Platinum Taq DNA Polymerase kit (Invitrogen, Carlsbad, CA, USA). In brief, the two reaction mixtures (25 μl) contained 12.5 μl reaction buffer (a buffer containing 0.4 mM of each dNTP, 3.2 mM MgSO4), 1 μl of enzyme solution (SuperScript III RT/Platinum Taq Mix), 1.25 μl of probe primers solution (10 µM stock concentration) 3 μl magnesium sulfate (50 nM), and 7.25 μl of RNA. The cycling conditions were as follows: cDNA synthesis at 55 °C for 30 min (min) and 50 °C for 2 min followed by 45 cycles of denaturation at 95 °C for 15 s (s), extension at 57 °C for 30 s and collecting the fluorescence signal at 68 °C for 30 s. Target 1 (E gene) forward primer ACAGGTACGTTAATAGTTAATAGCGT; reverse primer TGTGTGCGTACTGCTGCAATAT; and probe 5′-FAM-ACACTAGCCATCCTTACTGCGCTTCG-TAMRA-3′. Target 2 (N gene) forward primer GGGGAACTTCTCCTGCTAGAAT; reverse primer CAGCTTGAGAGCAAAATGTCTG; and probe 5′-FAM-TTGCTGCTGCTTGACAGATT-TAMRA-3′. As positive controls, double stranded DNA fragments (gBlocks, IDT, Belgium) with a five-nucleotide deletion in the targeted part of the E (102 copies/µl) and N (103 copies/µl) gene were used. Positive control Ct vales were 31.67 ± 0.68 and 28.07 ± 2.66 respectively. All PCR products with a Ct value < 45 were confirmed by Sanger sequencing (Macrogen, the Netherlands). Negative controls (swabs) were performed on non-exposed VTM (Suppl. Table 1).
Inoculation
Vero E6 cells (green monkey kidney cells (ATCC CRL-1586)) were seeded into T-25 flasks and grown in DMEM (Gibco, 41966) supplemented with 10% FBS (Gibco, USA) and 1 × Penicillin–Streptomycin (Sigma-Aldrich, PA333). The flasks were incubated (37 °C, 5% CO2) until cells confluency reached approximately 90%, after which the cell media was substituted with 9 ml of pooled samples supplemented with 2% FBS and 1 × Penicillin–Streptomycin. Potential cytopathic effect (CPE) was observed daily. Increase in viral load was determined by rRT-PCR, using 100 μl of supernatant from each T-25 flask at 0 (base line for comparison), 24 and 120 h post infection (hpi). rRT-PCR was also performed on DMEM exposed to air in a BSL-2 laboratory for 24 h (see section “Fluid samples”), non-exposed DMEM and DMEM spiked with SARS-CoV-2 synthetic oligonucleotide control sequence as negative and positive controls, respectively (Suppl. Table 1). Eleven days post inoculation, supernatants from the pooled samples (1 ml) were passed once into new flasks seeded with Vero E6 cells and containing 4 ml of cell media. Two days after the passage, samples were taken as described above for quantification by rRT-PCR. All procedures involving live virus were performed in a BSL-3 laboratory.
Ethical approval
Approval for accessing patient information was granted from the Swedish Ethical Review Authority DNR 2020-01787. As this retrospective data collection was considered completely anonymized by the Ethics committee, the need for patient consent was waived by the Swedish Ethical Review Authority. The study was conducted according to good clinical and scientific practices and following the ethical principles of the Declaration of Helsinki.